
Abstract
ARISING FROM X. Zheng et al. Nature 495, 507–511(2013)10.1038/nature11985
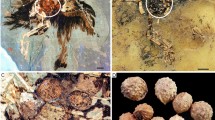
In a recent report Zheng et al. describe ovarian follicles in three fossil birds from the Early Cretaceous period of China belonging to Jeholornis and two enantiornithine species1. Because these were situated in the left half of the body cavity of the fossils, the authors suppose that the right ovary was already reduced in these early birds1. Fossilization of ovarian follicles would constitute an extraordinary case of soft tissue preservation, but the morphology of the fossil structures does not agree with the ovulation mode of coelurosaurs. There is a Reply to this Brief Communication Arising by O'Connor, J., Zheng, X. & Zhou, Z. Nature 499, http://dx.doi.org/10.1038/nature12368 (2013).
Similar content being viewed by others

Main
The Liaoning lagerstätten are renowned for many exceptional examples of soft tissue preservation in tetrapods2. However, integument preservation is usually due to fossilization of melanosomes2,3, and unambiguous evidence for the preservation of less resistible, melanosome-free tissue, such as muscles or internal organs, is scarce (note that the liver, which is sometimes preserved in fossils, contains a high amount of melanosomes). Although fossilized muscle fibres and gastrointestinal tracts of dinosaurs were reported4, some records, such as that of a supposed dinosaur heart5, were quickly refuted6.
In any case, the isolated preservation of easily perishable internal organs without fossilization of more durable soft-tissue structures, such as muscles or integumentary appendices, would be remarkable. In fact, two of the specimens reported by Zheng et al.1 do not show any traces of feathers, and specimen STM29-8 became fossilized in an advanced state of decay, with bones of the pectoral girdle being disarticulated. As can be observed in dissections of decomposed avian carcasses, the gonads are among the first visceral organs to fall victim to decay. Thus, it would be highly unexpected if follicles were the only preserved soft tissue structures. The assumption of Zheng et al. that mature follicles could have been preserved owing to fossilization of the “perivitelline layer and other protective layers”1 is not well founded, because in birds this layer consists of glycoproteins7, which are unlikely to fossilize.
The presence of up to 12 or 20 equal-sized mature follicles in the specimens reported by Zheng et al. would suggest simultaneous ovulation of many follicles, as in crocodiles. However, there exists evidence for paired shelled eggs in compsognathids8 and oviraptorosaurs9, and the eggs are arranged in pairs in the nests of oviraptorosaurs and troodontids10. This indicates that the avian ovulation mode, that is, the consecutive maturing of follicles, was already present in coelurosaurs, although these still retained two functional ovaries11. As a consequence, distinct size differences would be expected among maturing follicles of early Cretaceous birds.
It is also remarkable that the diameter of the largest “follicles”, 8.8 mm, is the same in all three specimens reported by Zheng et al., despite the fact that these animals differ greatly in size. We further note that interpretation of similar-sized, spherical structures in the holotype of Compsognathus from the Solnhofen limestone as eggs is likewise disputed12,13.
Although gingko ovules from Liaoning have a similar shape and size14, we agree with Zheng et al. that the morphology of the spherical structures in the bird fossils do not conform with those of ‘seeds’ (that is, fruit stones). However, in addition to fruit stones there existed other objects in Cretaceous ecosystems that could have been ingested by these birds, such as the fleshy arils of gymnosperms. Fossilization of such organic material in the acidic milieu of the stomach seems more likely than a selective preservation of soft tissue within the body cavity15.
References
Zheng, X. et al. Preservation of ovarian follicles reveals early evolution of avian reproductive behaviour. Nature 495, 507–511 (2013)
Benton, M. J., Zhonghe, Z., Orr, P. J., Fucheng, Z. & Kearns, S. L. The remarkable fossils from the Early Cretaceous Jehol Biota of China and how they have changed our knowledge of Mesozoic life. Proc. Geol. Assoc. 119, 209–228 (2008)
Zhang, F. et al. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature 463, 1075–1078 (2010)
Dal Sasso, C. & Signore, M. Exceptional soft tissue preservation in a theropod dinosaur from Italy. Nature 392, 383–387 (1998)
Fisher, P. E. et al. Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. Science 288, 503–505 (2000)
Cleland, T. P., Stoskopf, M. K. & Schweitzer, M. H. Histological, chemical and morphological reexamination of the “heart” of a small Late Cretaceous Thescelosaurus . Naturwissenschaften 98, 203–211 (2011)
Sasanami, T., Pan, J. & Mori, M. Expression of perivitelline membrane glycoprotein ZP1 in the liver of Japanese quail (Coturnix japonica) after in vivo treatment with diethylstilbestrol. J. Steroid Biochem. Mol. Biol. 84, 109–116 (2003)
Chen, P.-J., Dong, Z.-M. & Zhen, S.-N. An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 391, 147–152 (1998)
Sato, T., Cheng, Y.-N., Wu, X.-C., Zelenitsky, D. K. & Hsiao, Y.-F. A pair of shelled eggs inside a female dinosaur. Science 308, 375 (2005)
Varricchio, D. J., Jackson, F., Borowski, J. J. & Horner, J. R. Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits. Nature 385, 247–250 (1997)
Zelenitsky, D. K. Reproductive traits of non-avian theropods. J. Paleont. Soc. Korea 22, 209–216 (2006)
Griffiths, P. The question of Compsognathus eggs. Rev. Paleobiol. Spec. Iss. 7, 85–94 (1993)
Reisdorf, A. G. & Wuttke, M. Re-evaluating Moodie’s opisthotonic-posture hypothesis in fossil vertebrates part I: reptiles — the taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany). Palaeobio . Palaeoenv. 92, 119–168 (2012)
Zheng, S. & Zhou, Z. A new Mesozoic Ginkgo from western Liaoning,China, and its evolutionary significance. Rev. Palaeobot. Palynol. 131, 91–103 (2004)
Lerosey-Aubril, R. et al. Controls on gut phosphatisation: The trilobites from the Weeks Formation Lagerstätte (Cambrian; Utah). PLoS ONE 7, e32934 (2012)
Author information
Authors and Affiliations
Contributions
G.M. and A.M. contributed equally to the manuscript.
Corresponding author
Ethics declarations
Competing interests
Declared none.
Rights and permissions
About this article
Cite this article
Mayr, G., Manegold, A. Can ovarian follicles fossilize?. Nature 499, E1 (2013). https://doi.org/10.1038/nature12367
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature12367
This article is cited by
-
Reanalysis of putative ovarian follicles suggests that Early Cretaceous birds were feeding not breeding
Scientific Reports (2020)
-
Confirmation of ovarian follicles in an enantiornithine (Aves) from the Jehol biota using soft tissue analyses
Communications Biology (2020)
-
Early evolution of the biological bird: perspectives from new fossil discoveries in China
Journal of Ornithology (2015)
-
Zheng et al. reply
Nature (2013)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
